Fallet
Person förlorar synen på vänstra ögat efter en gallstensoperation (oklart om det är relaterat i skrivande stund). Undersökningar genomförs och Carotis Interna har plack. Pupillreflex fungerar inte heller på vänstra öget. Får acetylsalicylsyra och det går tillbaka. Lite senare får patienten bortfall på båda ögonen, nedre högra hörn. Synen återkommer sen igen.
Fallet inkluderar inte lukt och smak men ingår i veckan ändå.
Studiemål
Ögat
- Anatomi
- Blodförsörjning och Innervation
- Tappar och stavar
- Från hornhinna till Cortex (synbanan)
- Transduktion
- Hur omvandlas ljus till elektriska signaler
- Reflexer
- Synfel
Lukt och Smak
- Kort liknande som för ögat.
Kursguiden
Lukt och smak
- Översiktligt lukt- och smaksinnenas anatomi och fysiologi, från receptorstimulering till
central informationsbearbetning.
- Mycket översiktligt, lukt- och smaksinnenas roll i styrningen av olika slags beteende, samt den kliniska betydelsen av rubbningar i lukt- och smaksinnenas funktion.
Syn
- Översiktligt ögats anatomiska uppbyggnad, kunna namnen på de olika ögonmusklerna samt innervationen av dessa, i stora drag kunna redogöra för hur de olika ögonmusklerna bidrar till ögats rörelser.
- De delar av ögats anatomi som är av direkt relevans för dess funktion som sinnesorgan. Detta omfattar uppbyggnad av, funktion hos och inbördes relation mellan strukturer involverade i ljusbrytning, ackommodation, ljustransduktion och afferent transmission av nervimpulser.
- Ögats optiska egenskaper samt den strukturella och funktionella bakgrunden till de vanligaste brytningsfelen i ögat och hur dessa åtgärdas. Brytningsfelen omfattar myopi, hyperopi, presbyopi och astigmatism.
- Näthinnans neuronala uppbyggnad och funktion, speciellt med avseende på ljustransduktion i ljusreceptorcellerna samt impulstransmission från ljusreceptorer till synnerv. Mot bakgrund av detta kunna redogöra för likheter och skillnader mellan stav- och tappsystemet vad gäller känslighet, upplösning samt regional distribution av ljusreceptorer.
- Olika reflexer som verkar på ögat, redogöra för dessa reflexers anatomiska underlag och funktion samt i relevanta fall deras kliniska betydelse och testning.
- Synbanornas funktionella anatomi från synnerv till synbark samt definiera de bortfall i synfältet som uppträder vid skador på olika delar av synbanorna.
- Översiktligt, med utgångspunkt från begreppet "receptivt fält", viktiga karakteristika hos informationsbearbetningen i synsystemet, särskilt med avseende på uppdelningen i flera funktionellt specialiserade parallella kanaler.
- Översiktligt funktionell lokalisation inom de områden av cerebrala cortex som bearbetar olika aspekter av synintrycket samt kunna diskutera de experimentella och kliniska fynd som ligger till grund för denna uppdelning.
Översiktligt, med utgångspunkt från begrepp som "trichromacy", "colour opponent processes" och "colour constancy" redogöra för färgseendets fysiologi och anatomi.
Tentafrågor
- Vilken av näthinnans fotoreceptor-typer har fotopigmentet rodopsin, och vad består detta rodopsin av?
- Beskriv kortfattat vad som händer med den elektriska potentialen i en fotoreceptorcell i näthinnan som träffas av ljus.
- Vilken effekt har detta på cellens neurotransmittorfrisättning? 1p
- Beskriv vidare konsekvenserna för nästa neuron i händelsekedjan om detta är en bipolar ON- cell. Utgå gärna ifrån termer som ökad/minskad excitation/inhibition och ökad/minskad signalering.
- Vilken är främsta fördelen med binokulär syn (dvs att man har två ögon)?
- När information fortleds från ögonen till primära syncortex och vidare till associationsareorna går den till en början via monokulära neuron, dvs neuron som bara erhåller information från ett öga. Var i hjärnan slutar den monokulära informationsbehandlingen och var börjar den binokulära?
Sammanfattning
Synen - Introduktion
Ögats anatomi
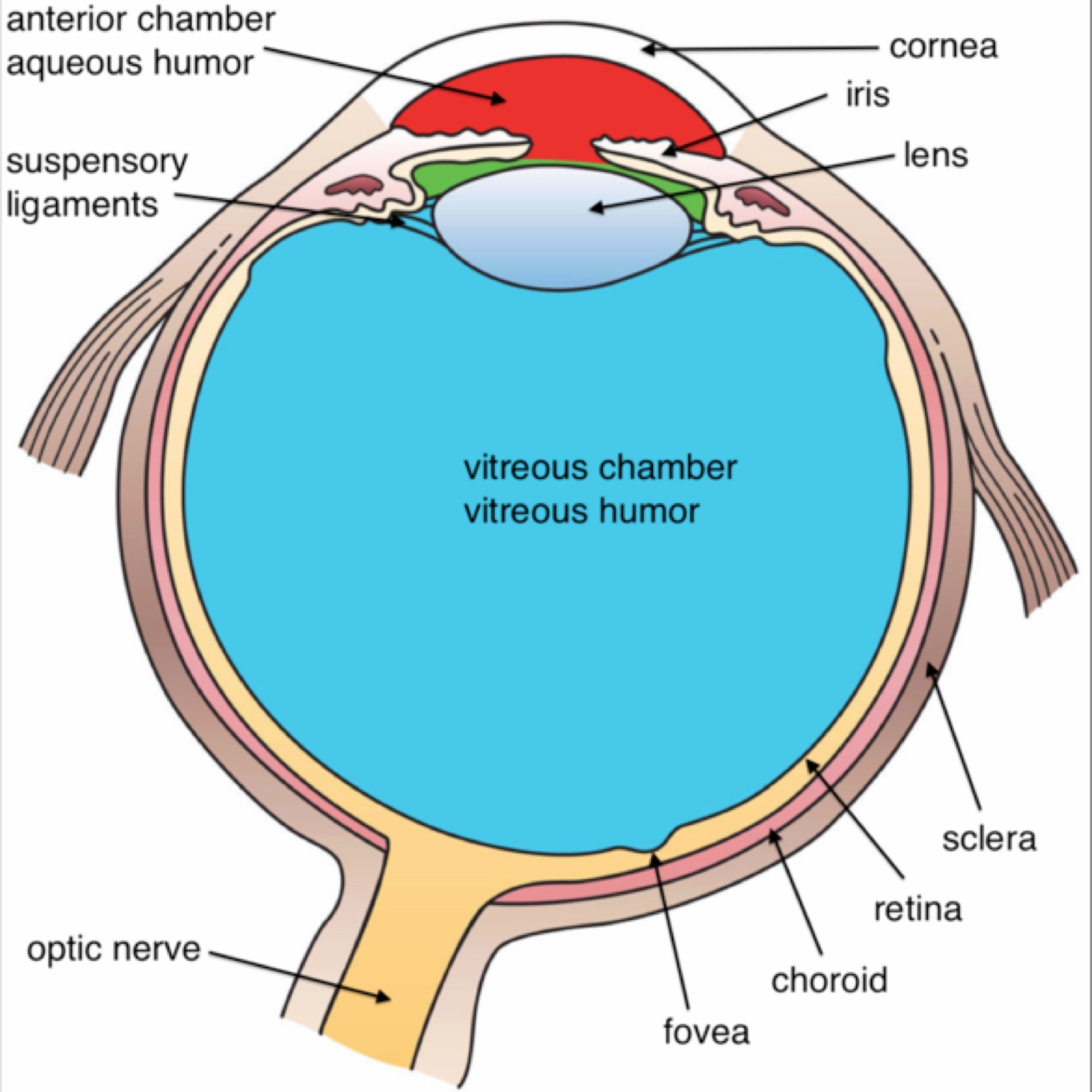
- Sclera: Vita hinnan
- Choroidea: Åderhinnan
- Retina: Ytan med stavar och tapar
- Fovea: Mest ljuskänsliga delen i Macula Lutea (gula fläcken), främst tappar.
- Macula Lutea: Området tätast med synnerver
- Discus Opticus: Där nerver och blid går in, vi ser inget (blinda fläcken).
- Cornea: Hornhinnan, första punkten att bryta ljus
- Pupilla: Öppningen för ljuset
- Lens: Bryter ljuset
- m. Sphincter pupillare: Sköter storleken av pupillen.
- N. Opticus: Sänder synsignalerna in i hjärnan.
- Tunica Conjuctiva: Smörjer ögat
- Retina: Där alla ljussensorer (stavar, tappar) sitter.
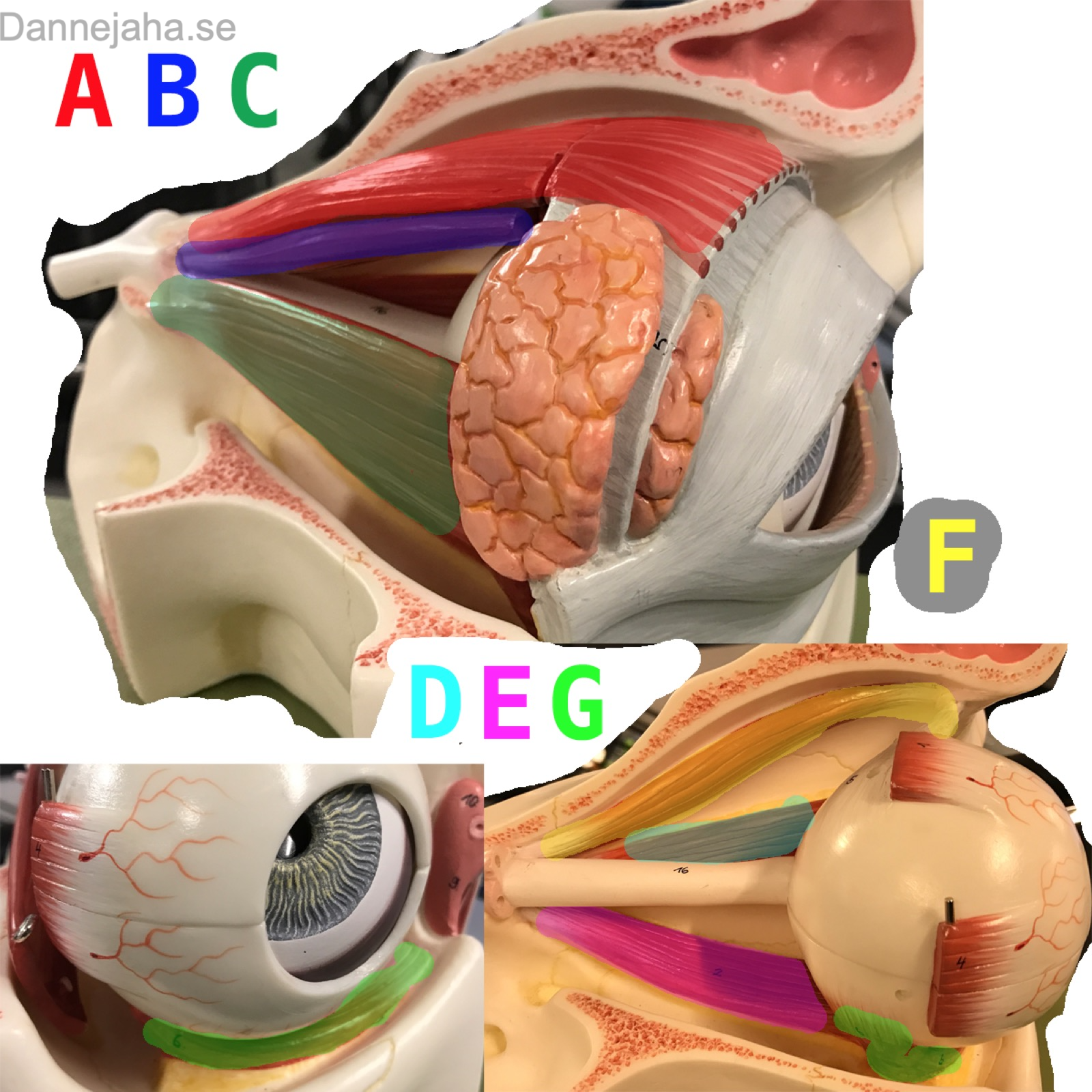
Muskler
- A: m. Levator Palpebrae Superiosis: Ögonlocket
- B: m. Rectus Superior: Uppåt
- C: m. Rectus Mediale: Innåt
- D: m. Rectus Laterale: Utåt
- E: m. Rectus Inferior: Neråt
- F: m. Obliquus Superior: Abducerar och roterar ögat
- G: m. Obliquus Inferior: Lyfta och Abducera ögat
- Stavar (rods): Ljuskänsliga och ser svart/vitt.
- Tappar (cones): Ljus och färgkänsliga (Röd, Grön, Blå)
- Ganglions: Bildar den n. Opticus.
- Bipolar cells: Processar och skickar vidare informationen från foto-receptorerna (stavar, tappa) till Ganglions.
- Melanin/epitel: Hindrar ljus från att reflekteras ut igen och störa synen. Ger näring till cellerna.
Notera att stavarna är vända bort från ljusets riktning och nerbäddade mot pigmentcellerna.
På grund av uppbyggnaden av hur en lins bryter ljus, blir det att vänster sida av ögat tar emot ljus från höger och taket tar emot ljus underifrån. Vi ser alltså upp och ner och spegelvänt. Hjärnan vänder rätt på detta för att ge en bättre representation av omvärlden.
Ögat består kort av 3 delar av lager med olika innehåll beroende på plats:
- Sclera och Chornea
- Choroidea, Iris, Corpus Cillare
- Retina (Stavar och tappar)
Bakre kammaren separeras från den främre av Lens. Inte att förväxla med Camera Anterior och Posterior som separeras av Iris. Främre kammaren får näring via vätska och dränerar sedan i Schlemm kanalen (Sinus Venosus Scerae).
Nervers innerverande på synen
- N. Opticus (2)
- Sensorisk för att ta emot synintryck
- N. Oculumotoris (3)
- M. Obliquus Inferior
- M. Rectus Superior
- M. Rectus Inferior
- M. Rectus Mediale
- M. Cilliares
- N. Trochlearis (4)
- N. Abducens (6)
Synintryck kopplas till fler ställen
Utöver då att vi uppfattar synen som en bild av något så kopplas även intrycken till andra delar av hjärnan för tolkning:
- Hypothalamus: Reglering av hjärtrythm
- Pretectum: Reflexkontroll av pupill och lins
- Superior Colliculus: Orientering av ögonrörelse i kombination med huvud.
Ljusets väg in i ögat
Lens och chornea är de delar som bryter ljuset (mest lens) och ser till att det träffar retina på rätt ställe och vi kan se skarpt. Lens styr med muskler för att ställe in avståndet, detta kallas Accomodation. Ju mer sfärisk formad Lens är, ju mer bryter den ljuset, alltså gör den att vi kan se saker närmare oss. Förmågan att göra detta blir sämre med åren och därför får vi svårare att se på nära håll. Musculus Cilliaris är de muskler som sköter denna biten.
När man slappnar av är linsen avslappnad (Sympatiska nerver) och det ljus som träffar ögat skarpt är det ljuset som syns på långt avstånd. Fokus ligger långt bort. För att se på närmare håll måste vi spänna muskel och dra ihop linsen (Parasympatiskt), mer och mer. Detta som äldre får svårare med då linsen blir stel, Ålderssynthet.
Eftersom vi har två ögon och får in bilateral information (alltså vänster och höger synfält från båda ögonen). Samtidigt som de skiljer sig något (sitter lite bredvid varandra), så får vi några fördelar.
- Vi kan mäta djupet
- Vi har en "backup" om något händer på ena ögat.
Man har dock ett dominant öga och detta kan testas genom att sätta fram handen, bilda en cirkel med tummen och pekfinger och "omringa" ett föremål långt borta. Blunda nu med ett öga åt gången. Om föremålet är kvar har du det dominanta ögat, försvinner det, har du det underordnade.
Ljuset som kommer in är alltså både spegelvänt (vänster/höger) men även upp och ner! Ögats fokus (det avlägsna) är runt 6 m bort. Efter denna punkt så kommer linsen vara av samma form oavsett avstånd vi fokuserar på. Detta beror på att ljus på länge avstånd i princip kommer in med samma brytning, parallellt. Närpunkten kan avgöras genom att ta ett finger och flytta närmare ögonen tills det inte längre är skarpt. Denna punkt flyttas längre bort med åldern.
Fotoreceptorer
I ögats retina finns två typ av receptorer, stavar (svart/vitt, främst i perferiet) och tappar (färg, främst i fovea). Dessa tar emot ljus och omvandlar det till signaler som sedan ska tolkas. Stavarna är mycket känsliga (så lite som 1 foton krävs), medan tapparna kräver mycket mer stimulans för att aktiveras. Därför har vi bättre seende i svart/vitt än vi har i färg. Både tappar och stavar är varierande känsliga för olika våglängder av elektromagnetisk strålning (ljus).
Att komma ihåg:
- Ljus hyperpolariserar fotoreceptorer
- Deoplariserade fotoreceptorer sänder ut signal!
Stavars mottagande av svartvitt
Retinal kommer från A-vitamin och är därför detta är viktigt för synen. Na+ kanaler är öppna om ljus inte stimulerar och cGMP finns tillgängligt (håller kanaler öppna). När ljus når stavarna kommer detta att leda till att Na+ kanalerna stänger sig.
- Ljus träffar Rhodopsin (Innehåller Retinal)
- Cis-Retinal ändras till Trans-retinal
- Transducin släpper då ifrån sig sin Alpha-enhet
- Detta får PDE att aktiveras och omvandlar cGMP till GMP
- GMP istället för cGMP gör att Na+ kanalerna stänger isg.
Signalen går vidare
Efter detta finns det ON- och OFF-Bipolära celler som tar emot denna stimulans.
- ON blir Inhiberade av Glutamat (ON-center av ljus, alltså närvaro av ljus)
- OFF blir exiterande av Glutamat (OFF-center av ljuset, alltså frånvaro av ljus)
Detta innebär att beroende på ljusintensiteten som träffar Stavarna (eller tapparna) så kommer ON eller OFF bipolära celler att vara mer eller mindre aktiva. Är det en Stav kommer den att koppla vidare via en Amacrine-cell innan den når Ganglion till n. Opticus. Tappar som förmedlar färg går direkt. Detta ger förmågan att uppfatta ett större spektrum av ljusintensitet.
Informationen som skickas till hjärnan är dock bara en relativ bild av det man ser. Signalering mellan cellerna, genom Horizontal cells, kommer de att kunna påverka aktiviteten hos stavar (mörker) och tappar (ljus). Det är detta som gör att ögat kan anpassa sig efter ljusstyrkan och även det som gör att en färg ser ljusare ut bredvid ett mörkare objekt. Anpassningen är lokal i ögat.
Horizontal cells stimuleras samtidigt som Amacrine-cells och ON/OFF-bioplar celler och när de stimuleras kommer de att utsöndra GABA som påverkar fotoreceptorerna intill. Detta för att öka kontrasten och därmed förmågan att se.
- Stavar: Tar emot svartvitt
- Tappar: Tar emot färg
- ON-Bipolära: Gillar ljus, ökar utsöndring vid mer ljus.
- OFF-Bipolära: Gillar Mörker, ökar utsöndring vid lägre ljus.
- Horizontal Cells: Ökar kontrast genom att ändra känslighet hos granncellerna
- Amacrine cells: Omkoppling mellan stavar och ganglion till n.opticus.
Tappars mottagande av färg
Tappar är mindre ljuskänsliga (1000 gånger) och är i stället för bara ljuskänsliga, färgkänsliga. Det finns tre typer; röd, grön blå. Istället för Rhodopsin så använder tappar Photopsin (Iodopsin är alternativt namn). Fördelningen mellan de olika färgerna är enligt nedan. Tapparnas responstid är också bättre än stavar, alltså de återställs snabbare.
Det är våglängdsomfånget för Photopsin som avgör viken färg eller våglängd de är känsliga för. Detta är ingen skarp exakt värde (typ 450 nmn) utan snarare en kurva och omfång med en topp vid en punkt. Alltså reagerar flera tappar på samma våglängd, som då ligger mellan deras toppar i spektrumet. Stavar ligger mellan röd/grönt spektrum (streckade linjen). Vitt ljus är egentligen en blandning av hela spektrumet, svart är frånvaro av ljus. Allt detta innebär att vi människor har Trichomatic syn. Det är på grund tappar med olika känslighet vi kan skilja på ljusets färg.
Det finns det människor med 4 olika typer av tappar (tetrachromacy), och färgblinda kan sakna någon med. Detta beror på att X-kromosomen har färgseende för röd och grön, och kvinnor har 2 X-kromosomer. Detta gör också det ovanligare för kvinnor att vara färgblinda än män. Det är dock ovanligt att man saknar 2 tappar, och än mer tre. Vanligast är att en saknas i så fall. Skador på retina eller V1 kan också orsakar färgblindhet.
Då tappar är snabbare än stavar kan det uppvisas under experimentet Pulfrichs Pendel. Man utsätter ena ögat för mörker och andra för ljus. Detta betyder att ena använder stavar och andra tappar för att se. Vad som händer då är att när man tittar på ett föremål som rör sig, är att stavarna halkar efter. Detta leder i sin tur att objektet ter sig skilja mellan ögonen och därmed uppfattas som att det finns längre bort i synfältet.
Från ögat till Cortex
Intrycket från ögat separeras och går till Nucleus Geniculus Lateralis och sedan delas upp sig för att gå till olika delar om Sulcus Calcarnicus. Den övre delen av ögat projicerar superiort om Sulcus Calcarnicus, medan den undre delen projicerar inferiort om den. För att nå inferiora delen i Area Striata måste man runda Ventriculus Lateralis i något som kallas Meyers Loop.
- Visuellt nere -> Taket i ögat -> Superiort om Sulcus Calcarnicus
- Visuellt uppe -> Golvet i ögat -> Meyers Loop -> Inferiort om Sulcus Calcarnicus
Meyers Loop är kliniskt viktig eftersom det är här det en av uppdelning av synnerverna sker och man kan se om övre/undre delen av ena synhalvan som tagit skada. Lokalisation av skadorna.
För att fördjupa sig ytterligare kan man se att Corpus Geniculum Laterale är uppdelad i flera lager (6st) som för signalerna vidare till Area Striata (som delas av Sulcus Calcarnicus). Dessa 6 lager delas upp så att 3 lager tar emot signal från ena ögat och 3 från andra. Signalen är alltså fortfarande uppdelad hela vägen fram till V1.
I Corpus Geniculum Laterale är även lagren uppdelade enligt Parva och Magno. Detta då M och P-celler från Retina föredrar att skicka signaler till vissa regioner inom Corpus Geniculum Laterale.
M-Celler
- Större celler
- Rörelse och kontrast
P-Celler
- Mindre celler
- Detaljer och färg
Väl framme i V1 kommer flera celler påverkas av båda ögonen, vara bioculara och detta utvecklas under barnets 3-7 första månader.
Genom flera anatomiska försök har man kunna påvisa att inte nog med att signalerna är strikt uppdelade och att man har en topologisk uppdelning, utan att synen verkligen är ett 1:1 förhållande mellan intryck till hjärna. Lite som pixlar i en datorskärm. Detta är både i Corpus Geniculum Laterale som i V1. Experiment med att lysa på delar av retina och mäta hjärnana aktivitet i Area Striata bekräftar också att så är fallet (djur). Hos människor har man undersökt personer med skador (t.ex. av skottdrama).
Vid andra försök (på människor) har man punktstimulerat delar av i Area Striata och då fått personen att uppleva ljuspunkter i synfältet, väl placerade efter var man stimulerar. Migrän eller epileptiska anfall som har sin gång i närheten av V1 kommer också ge vågmönster eller blixtrar.
Tolkning i V1 (Primära Syncortex)
Olika celler i V1 tolkar informationen olika och används därför för olika saker. Man har sett att det kan används för att tolka konturer, riktning av rörelse och även djupet. En del celler aktiveras bara om intrycket är lika från båda ögonen (biocular cells). Detta är dem alltid då ögonen har något olika placering, förutom just för center av Macula.
Typer av uppdelningar i Area Striata (Modular Organization) kan vara våglängd (färg), orientering på objektet eller vilket öga som är dominant och därmed störst inflytande.
Våglängdspecificitet verkar vara, till skillnad från andra oganisationer, ihopklumpade som blobbar snarare än kolumner i Lamina 2-3 i Area Striata.
Trots att allt verkar separerat så finns där inga tecken på att det finns någon tydlig slutdestination för alla intryck (hastighet, form, färg osv). Inte ens i frontalloben är dessa integrerade utan separerade. Hur vet vi då att bilen är gul och rör sig mot en? Var skulle denna sammansättning skickas (bilden) och vem skulle "titta" på den? Antagligen har vi informationen uppdelad och tolkar den.
Man har även gjort experiment med människor som saknar förmågan att se, och bett dem att fantisera ett objekt, säg en röd bil. Denna aktiverar då samma områden som skulle aktiveras vid seende, utom då V1. Man aktiverar de extrastriata delarna. Detta tyder på att man inte behöver V1 för att föreställa sig en bild från minne eller fantasi.
Avståndsbedömning
För att kunna avgöra avståndet använder vi flera mekanismer, en del är inlärda (3-6)
- Diskrepans mellan näthinnans mottagna bild. Större skillnad innebär att objektet befinner sig närmare.
- Konvergens. Närmare objekt, mer konvegens vid fokus på det.
- Perspektiv (vad finns i omgivningen)
- Färgspektrumförskjutning. Avlägsna föremål blåförskjuts
- Storleken på näthinnan
- Relativ rörelse. Objekt längre bort rör sig långsammare.
Alternativa vägar
Om man får skador så att information via Corpus Geniculum Laterale inte kan nå V1 kan man ändå få information genom en alternativ väg "Nucleus Pulvinaris" (del av thalamus) och "Superior Colliculus". Dessa har också bra mappning från retina men ger inte ett exakt synintryck. Dessa verkar istället ge information om rörelse och man kan därför registrera att något rör sig, men i övrigt är man blind. Personerna har alltså en känsla av att något rör sig, och i vilken riktning, men kan inte se det. Blindsynt!
Primära syncortex till Extrastriala areor
Det mesta går in i V1 och sedan vidare för processa i extrastriata visuella areor. Dessa hjälper till att tolka färg, rörelse och form. Detta har betydelse för om skador uppstår kan man har problem att uppfatta rörelse, även om man kan se. Det finns många sådana och man har hittat 10 i människor och 30 i apor. Antagligen har människan fler. Exakt hur allt fungerar är väldigt komplext och man vet inte helt.
För att få en klart bild av vad som händer visuellt måste V1 använda sig av de andra areorna i hjärnan. Den tar emot och tolkar till viss del men behöver form, djup, rörelse och färg.
Denna vidare bearbetning, skiljer från Retina till V1 med att här kopplas P och M-celler samman för att bilda intryck, snarare än att separeras som på vägen till V1.
- Vad? (Objekt identifiering)
- Ventral ström: Occipital Lobe -> Temporal Lobe
- Skador ger problem att urskilja kub från triangel
- Var? (Spatiala mönster och rörelse)
- Dorsal ström: Parietal Lobe
- Skador gör att man har problem att hitta saker i rummet och får alltså problem med målinriktade rörelser.
Tre primära vägar ut från V1 har identifierats och de hålls separerade till en början åtminstone och skickar vidare till V2 som ligger intill. M och P-celler är ganglion celler från retina. Båda varianterna finns som ON och OFF-celler.
- M-cells-lager
- Från Corpus Geniculum Laterale och verkar ge information om rörelse och djup.
- P-cells-lager
- Från Corpus Geniculum Laterale som ger information om form och mönster
- Våglängdsspecifika P-cellslager i Area Striata.
- Som ger information om färg.
Rörelse går vidare från V1 -> V2 -> V5 (Middle Temporal Visual Area). Det mesta härifrån går sedan till Posterior Parietal Cortex. - Dorsala vägen.
Färg går vidare från V1 -> V2 -> V4. Ventrala vägen.
Skador på synbanan
Efter att ögat har tagit emot de elektromagnetiska signalerna och tolkat om det så att n. Opticus ska skicka det vidare, kommer det att gå.
- Oculus
- N. Opticus
- Chiasma Opticus
- Corpus Geniculatum Laterale - Synapsar.
- Sulcus Calcarnicus - Synapsar igen
På vägen från ögat ner till Sulcus Calcarnicus kommer halva synfältet att korsa över i chiasma opticus för att låta den temporala vyn från ena ögat, möta den nasala av andra. Detta innebär att det vänstra synfältet från båda ögonen går mot den högra hjärnhalvan och tvärtom.
Man får tänka på att eftersom ljust bryts i ögat och det som finns på höger sida kommer in i vänstra delen av ögat (Temporalt)
Beroende på var man får en skada på nerven längs dess väg upplevs därför olika problem med synen.
- Left/Right Anopia (A):
- Blind på det ögat, precis vid ögat.
- Bitemporal Hemianopia (B)
- Rakt över Chiasma Opticus
- Blind på temporala delarna från båda ögonen (lateralt)
- Right Homonymous Hemianopia (C)
- Vid Corpus Geniculatum Laterale
- Högre/vänstra fältet på båda ögonen.
- Right Homonymous Superior Quadrantanopia (D)
- En del av axon på väg från Corpus Geniculatum Laterale till Sulucs Calcarnicus
- Delar av synfältet, t.ex. Övre högre på båda ögonen.
Skador kan även uppkomma på Macula (del i Fovea, gula fläcken) som gör att vi ser skarpt på den punkt vi fokuserar på. Detta kan resultera i förlust av skarpt seende (Macula skadad), Central Scotoma. Man kan även få skador på allt utanför i Retina (t.ex. vid Glaukoma) och då bara Macula finns kvar. Man får då tunnelseende (Constricted Visual Fields).
Ljusreflexer
När man lyser in i ena ögat kommer båda ögonen att reagera med ljusreflex, så att säga konsensuellt svar. Singnalerna går enligt flödet nedan.
- Ljus kommer in genom pupillen in till Retins och skickas vidare av n.Opticus
- Vid Chiasma Opticus kommer signalerna från ena halva av Retina att byta över till andra hjärnhalvan och andra stanna kvar.
- Vid Corpus Geniculum Laterale (på båda sidor), kommer dels synaps till V1 ske men även axon gå vidare till Nucleus Pretectal och synapsa där.
- Vid Nucleus Pretectal kommer ytterligare synaps ske och här kommer signal skickas till båda hjärnhalvors Nucleus Edinger Westphal (som styr n. Oculomotorius)
- N. Oculomotorius kommer nu att ställa in pupillen på respektive öga efter ljusflödet genom att stimulera musculus cilliaris.
Skador
Detta innebär att vid skada bakom ena ögat, kommer ljusreflexen inte att fungera från det ögat (ingen av ögonen reagerar). Däremot kan ögats pupill reagera om man lyser i motsatta ögat, förutsatt att n. Oculumotoris inte också är påverkad.
Får man en skada vid Chiasma Opticus, så att synapser inte når över till andra sidan, kommer man fortfarande få reflexsvar. Detta då överkorsningen inte sker här utan först efter Corpus Geniculum Laterale, och in i Nucleus Pretectal.
Vid skada på n. Oculumotoris på motsatt sida man belyser, kommer pupillen inte att kunna regaera, däremot kommer ljusreflexen fortfarande kunna ske.
Sjukdomar, problem och tillstånd
Color Constancy
När man genom att titta på ett föremål i olika ljus ändå kan avgöra vilken färg den har. Vi verkar göra detta genom att jämföra med omgivningen och på så sätt kunna säga att objektet är av en viss färg.
Afterimages
När man stirrar på en monoton bild, säg röd, så kommer man sedan när man byter och tittar på en vit bild att se grönt. Detta beror på att känsligheten hos de röda tapparna sänkt och nu dominerar då de gröna. Samma sker mellan gult och blå färg. Det är dock mer komplext eftersom även dessa typer av bilder har med Color Constancy att göra och inte bara våglängd. Alltså sker även tolkning senare.
Myopi- Närsynthet
Detta innebär att fokus kommer hamna framför fovea på retina och därmed börja dela sig igen och bilden blir osynlig på långt håll. Man ser på nära håll. Man åtgärdar detta med linser eller glasögon som är konkava, öppnar upp ljuset så att fokus hamnar rätt.
Hyperopi - Översynthet
Personer med detta tillstånd kan se föremål långt borta väldigt klart. Detta beror på att ljuset som kommer in böjs, men inte tillräckligt. Detta gör att fokus-punkten hamnar bakom fovea och därmed blir bilden inte skarp.
Lösningen blir att sätta en konvex lins framför cornea (hornhinnan) så att ljuset konvergerar och fokus hamnar i fovea. Med ålder blir dock musklerna som styr cillierna än mer förslappade, vilket innebär att man kommer behöva öka styrkan på linsen.
Presbyopi - Ålderssynthet
Beror på att linsen blir för stel med åldern och därmed förlorar förmåga att ändra form och ställa om sig för att fokusera. Man kan avhjälpa detta genom att byta ut linsen mot en ny artificiell lins.
Astigmatism
Innebär att cornea (hornhinnan), som är den första brytande delen (lens andra) är oregelbunden, vilket leder till att man får flera fokuspunkter, ljuset sprids. Detta leder till suddig bild. Kan hjälpas med hjälp av linser.
Glaukoma - Grön starr
Har man ett stegrande tryck i ögat, som beror på sämre bortförsel av kammarvätska, kommer trycket mot ögat att öka, retina. Detta kan leda till bortfall och celldöd, vilket slutar med att man förlorar delar av synen. Det kommer krypande utifrån och in och kanske inte märks direkt. Man gör tryckundersökningar med hjälp av kon som placeras mot ögat.
Cataract - Grå starr
När ljus inte kan ta sig in i ögat pga fibrernra i linsen har blivit grumliga. Får små barn detta och det inte åtgärdas snabbt kan permanenta skador av synen uppstå. Man brukar operera det genom att ta sönder linsen med Ultraljud, suga ut den och ersätta med ny syntetisk.
Antons Syndrome
När man fått skador på de occipitala kortikala delarna i hjärnana. Patienten påstår att den kan se, och får saker presenterade för sig som inte finns.
Lukt och smak
Till skillnad från synen som använda fotoreceptorer och känsel som är mekanisk, så använder smak och lukt kemiska receptorer.
Lukt
För att man ska kunna lukta något så måste man få in molekyler från det ämnet. Det räcker med relativt små mängder, men det måste ske. Detta gör det intressant med just pengar (mynt) som luktar, det har alla känt. Detta beror inte på att metallerna löser sig i luften, utan på att det fett vi har på huden oxideras på metallen och i sin tur luktar. Mynt luktar inte. Våra 10 miljoner receptor celler hjälper oss att identifiera ungefär 400,000 olika lukter. Dessa neuron dör och ersätts, vilket är ett undantag till regeln att neuron inte kan ersättas.
Man ska också veta att lukt står för stor del (80%) av smaken. Detta visar varför man smakar dåligt vid förkylning och även varför smaken är dålig uppe i ett flygplan, där luften är dålig. Detta är möjligt eftersom vi har en passage i svalgen som kopplar ihop näsa och mun, där kommer partiklar gå upp. Man kan med hjälp av tungan smaka de olika kategorierna (salt, surt, sött, beskt, umami) men inte avgöra om det ät t.ex. citron eller lime.
En gammal myt är också att de olika smakerna registreras i olika delar av munnen, det stämmer inte. Alla smaker finns över hela tungan.
I nästans övre vägg finns små hålrum i benet (Lamina Cribrosa) som våra luktceller går igenom för att ta emot signaler från näshålan. Dessa receptorer är känsliga för specifika ämnen, molekylformer. Detta kopplar sedan vidare till Glomerulus celler och sedan Mitral och Tufed Cells för att sedan gå till upp i hjärnan.
- Kemisk partikel
- Receptor celler i Olfactory epitel
- Glomerulus (Garnboll)
- Projektionsnueron Mitral och Tufte Cells.
En viktig och unik del med lukten är att den till skillnad från andra sinnen går direkt från sinnescellerna till luktcentra i hjärnan, utan att tolkas eller synapsa via Thalamus. Alltså får vi in en ofiltrerad version av lukten till cortex. Detta kan delvis bero på att lukt är vårt äldsta sinne och tidigare i evolutionen stor del av vårt intryck, nästan hela cerebrum.
Luktens väg till hjärnan
Våra receptorer (Odorant Receptor Proteins, Ors) är inte specifika för vissa ämnen utan istället för visa formationer och isomerer. Liknande luktar snarlikt. Detta beror på att proteinerna som känner av är känsliga för form snarare än exakt molekyl. Anledningen till att vi kan känna sådan variation är att en doft kan binda till flera receptorer med olika delar av sin struktur. En kombination av detta blir som en tavla som ritas upp med doften.
När en doftmolekyl träffar en receptor kommer G-protein att aktiveras, cAMP och Na+ strömma in, en depolarisation av cellen.
Lukten fungerar delvis som synen och Olfactory Bulp är som retina, en mindre "hjärna" som är utanför den egentliga. Denna processar informationen som kommer in från de olika sensorerna. Luktsinnets Granule cells har dessutom också lateral inhibitation, precis som synen har.
Lukten terminerar till flera ställen men främst Uncus, Amygdala och temporalloben.
- Septal Nuclei
- Hypothalamus (Aptit, matsmältning, födobeteende)
- Får även information från bland annat Orbitofrontala cortex. Alltså får den både färsk information och bearbetad.
- Uncus -> Orbitofrontala Cortex
- Amygdala -> Mediodorsal Thalmatic Nucleus -> Orbitofrontal Cortex
- Olfactory Nucleus
- Finns på vardera sida och är sammankopplade (Liknande Chiasma Opticus). Alltså får båda hemisfärer information om lukt från båda näsborrar.
Det finns två olika typer av celler som aktiveras olika snabb, av olika mängder och olika specificiteter.
- Mitral: Skapa någon typ av kontext av doften
- Tufted: Snabbt aktiveras för att avgöra fara
Luktens topologiska indelning är inte alls så specifik som den är för t.ex. synen och känsel, där det sker väldigt bra mappning. Ute vid Glomerulus celler som tar emot luktsignalerna sker viss uppdelning. Dessa har en specificitet och tar emot signaler från liknande mottagceller (receptorer). Detta verkar dock inövat och vid tidigare stadier (hos möss) så tar Glomerulus cellerna emot signal från mottagarceller med olika specificiteter.
Skador på Unculus
Får man skador på Unculus eller dess omgivning kan det ge en känsla av doft (ofta ingen god) som ger en känsla av att man känt den förr (deja vu). Detta kan sedan leda till epileptiskt anfall och är en form av epilepsi.
Signaler från Primära Luktcortex
Signaler från Amygdala och Primära luktcortex går vidare i hjärnan för mer tolkning och accossiationer. Den fortsätter till Frontalloben (Orbitofrontala cortex). Orbitofrontala cortex kan även få signaler indirekt från Amygdala via Mediodorsala Thalmatiska kärnan. I Orbitofrontala cortex samlas information om känsel, minne, smak, känslor, motivation osv.
Feromoner
Omtalat ämne där djur använder feromoner för att hitta sexuella partners eller känna igen saker. Detta ska styras i Vomeronasal Organ, som verkar försvinna innan födsel hos människor.
Man har dock gjort undersökningar med kvinnors menstruationscykel, där det tydligen ska synkas om de lever ihop. Detta står i Brodal och Purves. Dock har detta betraktats snarare som seglivad myt eftersom det inte är något gäller generellt och flera studier har misslyckats med att reproducera det. The Guardian skrev 2016 Aug om detta och även professorer i Lund har nämnt på sina föreläsningar i organisk kemi.
Där finns dock andra tecken på att vi påverkas av dofter, som att man i väntrum penslat ämnen under stolar och sett hur beteendemönster för kvinnor och män i rummet ändrat sig.
"McClintock's original study (
McClintock, 1971) reported that onsets for dormitory friends became 2 days closer together over a 4–6 month period. Calling this result `menstrual synchrony' she encouraged the impression of menstrual concordance. In later critiques, Wilson (1991, 1992) noted three statistical errors in the evidence for menstrual synchrony: (i) failure to control adequately for the convergence of onsets by chance; (ii) inflation of the initial difference in onsets resulting in the spurious conclusion of synchronization over time; and (iii) sampling biases. Studies that correct for these statistical errors have found no evidence for synchronization, even as a weak effect (
Jarett, 1984;
Wilson et al., 1991;
Trevathan et al., 1993).
"
Tryffel
Grisar använder sig av feromoner för att veta om de är redo att para sig. I tryffel, så finns ett ämne som är just en könsferomon hos grisar och det är därför de är så duktiga på att leta upp dessa. Välj dock rätt kön på grisen... annars fungerar det inte.
Smak
För att känna smak måste molekyler binda in till tungas receptorceller och matcha. Det finns en intressant ämne "Miraculin" som kommer från mirakelbären. Detta ämne binder till söta receptorer men triggar inte dem. Det är först när man tillföra surt i munnen och molekylen i Mirakulin omformar sig, som de söta receptorerna sänder signal. Detta betyder att om man äter Miraculin, kommer surt att smaka sött, mycket sött. Detta är ett protein och bryts ner efter en kort stund.
Den sista och nyare smaken (Umami) triggas av glutamat och är också därför E621, smakförstärkare, eller MSG används i maten. I Asien är det vanligt med MSG för att få mer smak.
Tungan är indelad i smak-regioner (dock inte som myten att man smakar en viss smak på ett visst ställe). Det som stämmer är att tungan är mer känslig för vissa smaker inom vissa regioner. Man kan smaka alla delar över hela tungan.
Smakupplevelse är väldigt speciellt och det krävs mer än bara smakreceptorer för att få en upplevelse. Det är både textur (känsel) och lukt inblandat. I själv verket är smaken inte den större delen av upplevelsen. Samma pommes frites är betydligt äckligare om de mjuka snarare än krispiga och om du är förkyld snarare än frisk (dålig lukt).
Smakreceptorer
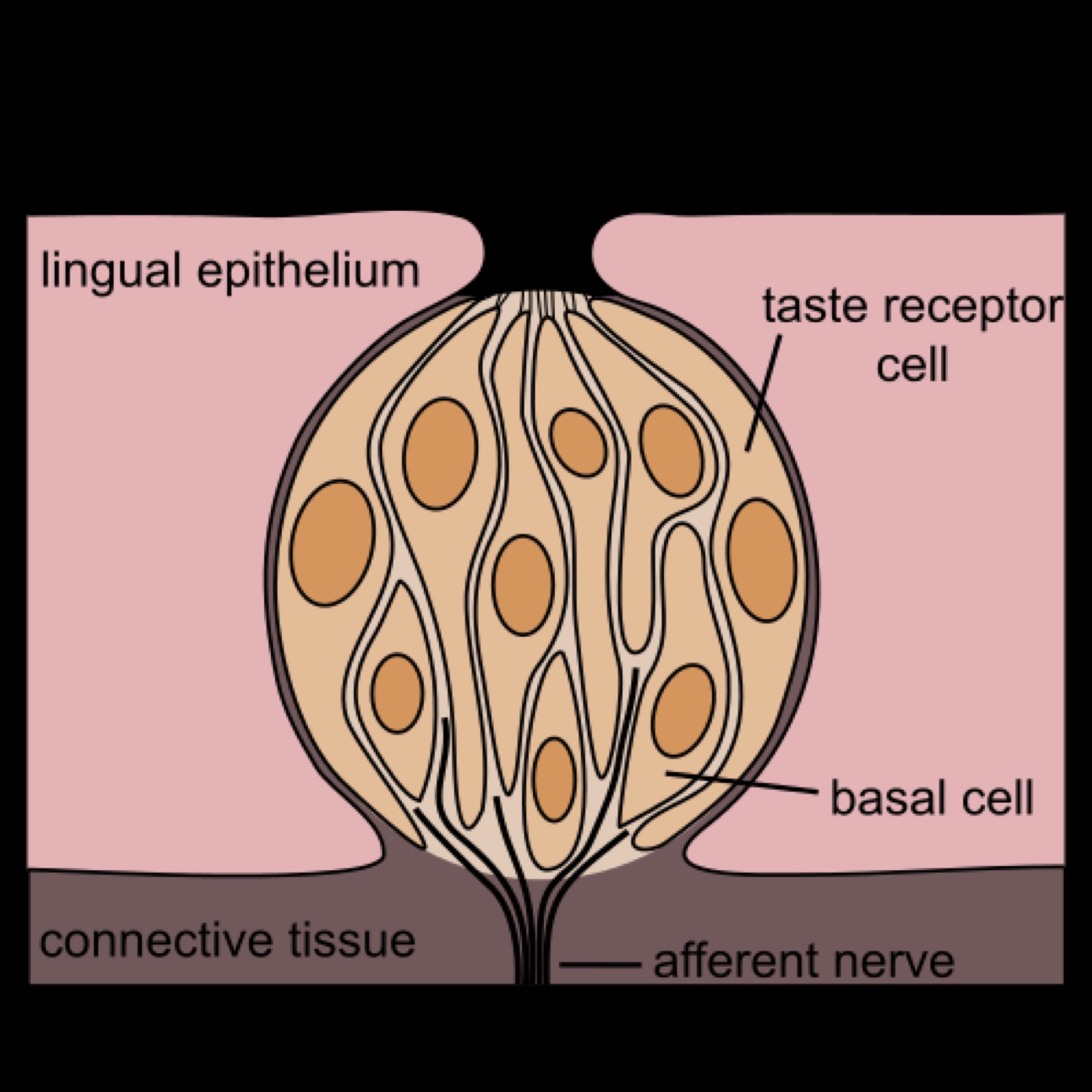
Sött, bitter och umami fungerar genom att aktivera G-protein kopplade receptorer. Surt och Salt däremot verkar svårare att förstå. Det kanske inte behöver specifika receptorer eftersom detta består av Protoner eller Na+ (H+ och Na+ joner).
Tolkning av smak
Smakkänsligheten kan variera genetiskt men verkar även bero på vår välmående. Serotonin är en signalsubstans som finns i våra smaklökar och vid depression eller ångest så minskar denna mängden och vår smakupplevelse blir sämre.
- Gustatory celler
- Basal-celler - Stamcellerna: Ersätter Gustatory-cellerna ungefär varje vecka
Smaklökars signal färdas:
- N. Facialis (intermediate, främre 2/3 av tungan) och n. Glossopharangeus (bakre 1/3 av tungan)
- Solitary Nucleus i Medulla Oblongata
- Hypothalamus: Påverkar högre autonoma center
Parabrachial Area i Pons -> Amygdala: Koppla smak till känslor
Thalamus: Skicka signaler till S1 nära där tungan representeras men även till Smakcentra i anteriora Insula som då blir det primära smakcentrat.
Sammankoppling av smak och lukt sker sedan i Orbitofrontal cortex (och några andra ställen).
Även Salivkörtel och parasympatic kärna av n. Vagus kopplas via Solitary Nucleus och detta leder till att matsmältningssystemet jobbar och att saliv utsöndras.
Smaksensorernas ledning är separerade tills de når Solitary Nucleus där de kopplas samman. Vissa svara dock enbart på sött eller bittert, men främst är det en blandning av signalerna man får. I Insula verkar det vara ytterligare mer sammankopplat. Men har kunnat se att där är viss urskiljning mellan de olika smakerna i cortex, dock inte helt fastställt.
Solitary Nucleus får signaler från andra delar när den ska tolka smak. Amygdala, Vagus (magen) och även blodet (glukos och insulinnivåer). Allt detta gör att smak kan förändras beroende på kontext, förväntningar och hunger. Detta kan ses bland annat genom att man ogillar fotsvett men kan tycka Port-de-salut ost är god. Trots det är samma molekyl som ger dess lukt!
Övrigt
Mycket är hämtat från Brodal - The Central Nervous System
Inspiration och populärvetenskapig
Föreläsning av Professor i Kemi; Ulf Ellervik som finns på LTH